
提取細菌外泌體的差異與相同要點
ISOLATION OF BACTERIAL EXTRACELLULAR VESICLES: WHAT'S DIFFERENT AND WHAT'S THE SAME?
提取細菌外泌體的差異與相同要點
關於細菌的細胞外囊泡 Bacterial extracellular vesicles (bEVs)之提取和分析, 需要對細菌生物學和欣賞如何與其已被深入研究的真核細胞外囊泡的對應物和bEVs之比較/對比有所了解; bEVs的研究仍然是一個新興的領域並帶有很多變數, 引發的事實是細胞外泌體可以被上百種不同的物種所製造出, 有些物種成分和EVs都可以被快速的轉變。
儘管如此, bEVs的研究卻是值得投入的, 因為bEVs代表的是一個非常重要的生物科技平台, 可在許多的醫療相關應用中顯示出無限的潛力。
細菌生物學的細胞外囊泡提取要點
細菌和古生菌屬於原核生物領域, 而細菌細胞的結構與真核細胞(動物中發現)有很大的不同; 雖然原核生物是沒有細胞核、細胞器或細胞骨架的單細胞生物, 可其卻內含有基因的物質(DNA, RNA)與生物分子合成、代謝和其他細胞活動所需的機能。
與真核生物相比, 原核細胞的尺寸更小其範圍大約在0.5-2 µm之間; 細菌的生長是很快速的, 當哺乳系細胞處於18-72小時的複製時間下時, 原核生物通常可在20分鐘內複製, 這可使其全生長培養(飽和的細胞)在8-10小時內完成。
這種快速的生長可使其在不斷變化的環境下生存, 代表細菌可經由複雜的基因表達調控網路來快速的適應; 雖然細菌有一般性的細胞骨架, 但其細胞壁卻有顯著的結構差異, 可使細菌被廣泛的分類群組如革蘭氏陽性、革蘭氏陰性或抗酸細菌。
重要的是, 這些細胞壁的差異可顯著的影響到細胞的膜所衍生出的奈米結構物製造 – 例如: 細胞外囊泡。
細菌外的外體之術語、成分和生物源
當第一次發現於革蘭氏陰性細菌中的bEVs時, 因其所被推測的細胞的來源, 當時就被歷史性的命名為”Outer Membrane Vesicles (OMVs)”- 外膜囊泡; 相反的是, 在最近被發現於革蘭氏陽性的bEVs被稱為是”Membrane Vesicles”- 膜囊泡, 其也使用了相同概念的細胞來源。
然而現在已知的是, bEVs不只存在於原核生物中也會於所有的生活領域中存在, 例如真菌、原生動物和植物細胞; 因此在本欄中, 我們都使用bEVs這個術語來泛指所有細胞所分泌的EVs, 這也是遵循了我們所述關於MISEV的指南要求的專欄。
bEVs是由親本細菌細胞所衍生出的脂質的奈米結構物(約25-300 nm), 其內含了多樣化的分子成分例如核酸(DNA, RNA)、蛋白質、脂質其他有機物質等; 更近的研究顯示, 在不同的生物發生機制下可以實現bEVs的製造, 其中某些EVs可能來自於起泡細胞, 而其他的EVs則是在細胞裂解的過程中被分泌出。
這些群體內的每個EVs都包含了從細胞的來源所帶的”貨物”: 於膜起泡型中的富含膜的貨物與在細胞裂解型中的富含膜/胞質的貨物; 在此方式下, bEV生物發生的主要機制是由於細胞對環境的反應調節的, 因此會影響bEV的成分。
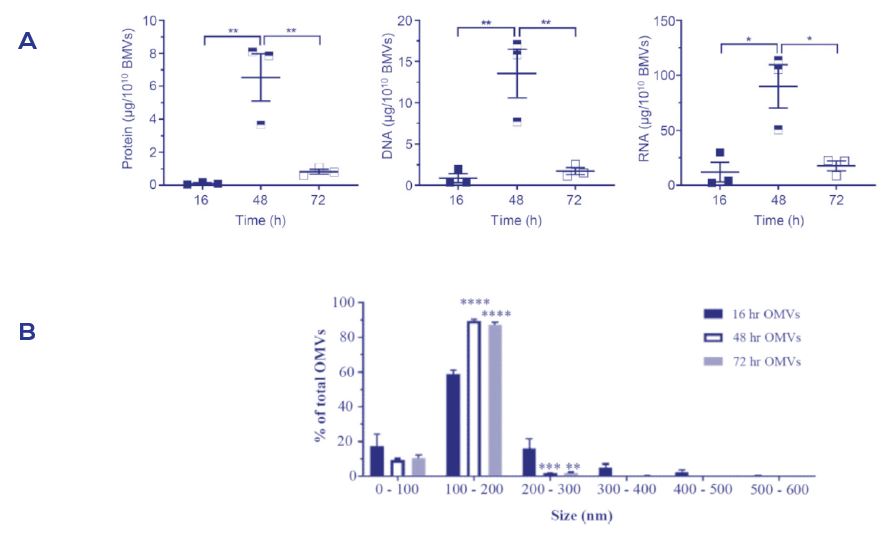
Figure 1. 由於培養條件導致的bEV成分的變異
取自幽門桿菌內的bEVs於第16小時(早期指數生長期)、第48小時(指數增長後期)和第72小時(靜態成長期)的蛋白、DNA和RNA數量(A)與粒子的粒徑分佈(B)的差異。
提取bEV前之變量的重要性
如之前所述, 細菌的快速生長可以產生對bEVs調查的顯著質疑性, 對環境線索的快速回應也引發了可調式-bEVs的貨物快速轉變, 以能夠應對新的條件或刺激, 這導致了異質性bEVs的分佈會被實驗條件依賴的因素嚴重的影響。
細菌培養的上清液條件可能有顯著的差異如下:
-
培養基 (富含、最小或生理性)
-
主要的細菌生長模式 (於生物膜或懸浮液的細胞生長)
-
生長階段 (每日或靜止)
-
溫度 (4-37 ℃範圍)
-
氧氣程度 (好氧、微氧或厭氧)
-
特定激發物的存在或移除
在此方法下, 有時候研究者不會察覺這些變數中的細微變化, 但會被細胞所引發的不同成份組成的bEVs所感知到; 在bEVs組成中, 培養條件的重要性已可開始被解決, 利用研究來表明不同的培養條件如何影響 bEV 組成, 進而在蛋白與RNA程度上進行更詳細的說明。
bEV貨物的廣義定量也顯示出其組成的變化是由於其培養條件(Figure 1A), 更多的是, 培養的變量可以影響bEVs的粒徑大小, 在下游的實驗中發現有陸續的影響; 這表示由不同的細菌生長階段下所收集到的bEVs表示了不同bEV粒徑的異質性(Figure 1B), 其被不同的宿主細胞攝取機制所優先主導。
因此當提取bEVs時, 強烈建議對bEV的製劑時的培養的變量進行徹底和全面的記錄, 以減少批次間的變異性並提高再現性。
如何得到”無細胞”的bEV樣品
懸浮生長的細胞(非生物膜所生長的細胞)所得到的細菌之細胞培養上清液(CCM)將會有大量的細胞, 一般從CCM中移除細胞的方法包含有離心、過濾或同時使用; 雖然細菌細胞可有不同的形狀, 但在單一細菌培養物中的細胞大小卻已知是異質性的。
多數用來將細菌的細胞顆粒下沉所用的離心法包含從4,000-10,000 g/10-20 分鐘的離心速度範圍; 然而, 依照由細胞製造的細胞外成分, 離心底部顆粒的一致性可更緊緻或鬆散, 這使完全達到無細胞的上清液轉移變得更困難與低效。
過濾法是利用將CCM通過微濾裝置而可處理不同的樣品體積, 或是由不同的離心力來推動樣品(例如注射器或幫浦過濾器); 過濾用的孔洞大小一般的範圍是0.2-0.4 µm, 這可有效的保留大多數的細菌細胞大小而不論其通過過濾器的形狀或角度(如縱向通過的棒狀細菌) – 這可使大多數的bEVs通過其濾液。
過濾法的最大限制是當去除細菌細胞時造成的濾膜堵塞, 原因就是細菌細胞與CCM所分泌的成分的富集。
因此, 將離心法與過濾法相結合可對整體細胞的耗竭有整體性的效率改進; 離心可移除主要的細胞並製造出澄清的上清液, 接著用過濾在最小的過濾膜堵塞概率下移除殘留的細胞。
提取bEV的方式
以細胞培養為底的模型是研究的主幹, 其可輕易的模擬許多不同生物體的細胞與生物過程; 自然的, 從生物流體中分泌的bEVs做為一個非常新穎的領域時, 從細胞培養中衍生的bEVs就成為主要的研究樣品。
與其他相似的懸浮性奈米結構物一樣, 細胞上清液的無細胞bEV樣品可由單一原理或多種原理結合的分離方式處理; 細菌細胞培養通常會在大體積的介質中進行(> 200 mL), 因此這通常都需要在使用適當的bEV分離方式前進行內含bEV上清液的”濃縮”。
bEV樣品的濃縮通常可由超濾的技術來達成, 其裝置有不同的容量、流體壓力或過濾孔大小, 我們推薦的超濾過濾器的孔洞大小是可以消除大部分污染性蛋白質, 同時保留大的奈米結構物/bEVs; 通常使用的是100 kDa的分子截流量(MWCO)過濾器, 而有時候MWCO大於300 kDa也能有效的處理。
超速離心(UC)為基礎的技術, 和哺乳類的EVs範例相同是最常被想到的bEV提取; 儘管如此, 在現有的文獻中並沒有對於UC的離心速度用於bEVs顆粒化或是bEV被顆粒化的分佈程度所形成的共識, 已發表的文獻其UC的離心速度範圍從40,000-400,000 g都有, 需要有更多的研究才能對於bEVs使用UC法的影響有更好的了解, 就像UC法對於哺乳動物EVs的功能有存在某些不利的影響一樣已被證明了。
密度梯度離心法(DGC)非常的耗時而冗長但卻被推崇為高純度的EV純化方式, 其利用了大小和浮力密度的差異來分離EVs和非EV的成分, DGC法已顯示出其在哺乳動物時可有效的分離EVs和非EV結構物, 像是富含在血獎內的極低密度與低密度脂蛋白; 然而這種方法依賴的分離效率無法轉移到bEV的領域內, 在缺乏於所有的bEV製劑中定義非EV的汙染物時, bEV的純度就無法被評估出。
現在bEV的研究中, 從特定的細菌菌株和生長條件的非EV之汙染物中能提供關於EVs提取方式的效率之觀點; 例如在兩種提取方式的bEV製劑中的蛋白組學分析提示了其可被用於當作汙染標記物的蛋白候選物, 其並非與實際的bEV相關, 可被以更強力的bEV提取方式移除。
在此方法下, 研究已顯示出存在於UC顆粒化(低純度法)的bEVs中的一些蛋白物, 再以DGC法(高純度法)再次進行後其豐度有明顯的下降; 或是已證實以DGC法可得到富含bEV純淨的標記物, 而非用UC法。
另一方面, 有了bEV組成中顯示的巨大可變性的支持下, 報導表示bEVs也會在一個、多個或任何密度階層與梯度中發現; 更多的是培養條件的單一變化就可引起DGC法的bEVs有不同的分佈模式, 這表明bEV的組成有發生了顯著的變化。
DGC階層可使用高分辨率的顯微技術而被目視到以確認EV的富集, 例如穿透式電子顯微鏡(TEM)、cryoTM冷凍電鏡或將已知的標記做免疫印跡; 然而有些研究顯示出DGC法的富含bEV的階層可因其高程度的總蛋白量和粒子計數而可被快速的分辨出。
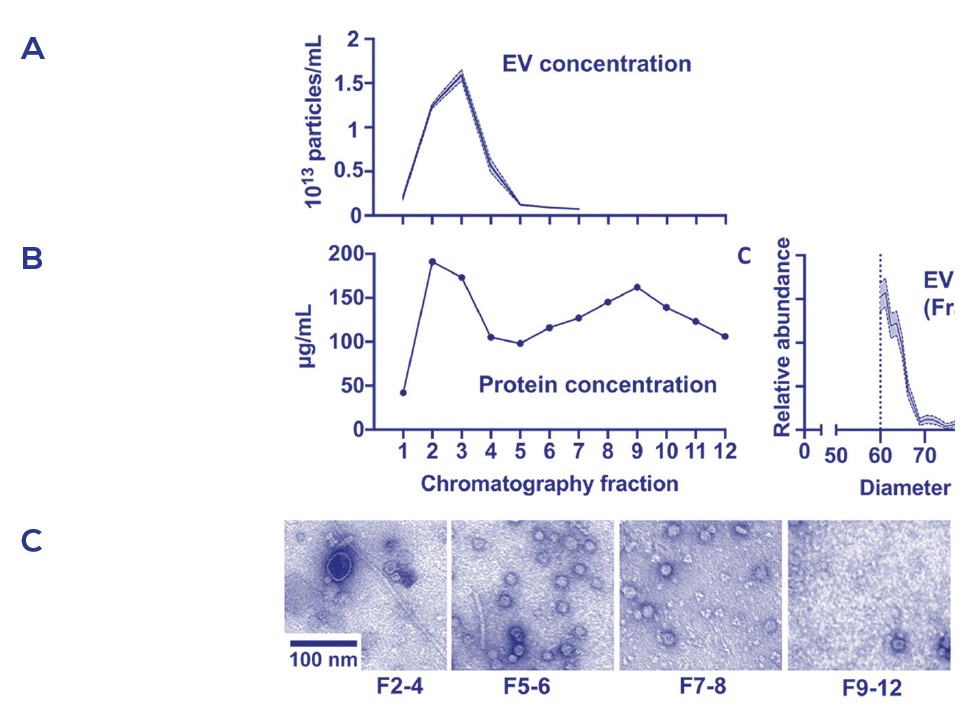
Figure 2. 由SEC的qEV columns純化的bEVs
使用35nm孔徑的qEV2管柱純化收集從大腸桿菌的bEVs的粒子濃度(A)和蛋白濃度(B), 收集體積的透射電子顯微鏡照片(C)。
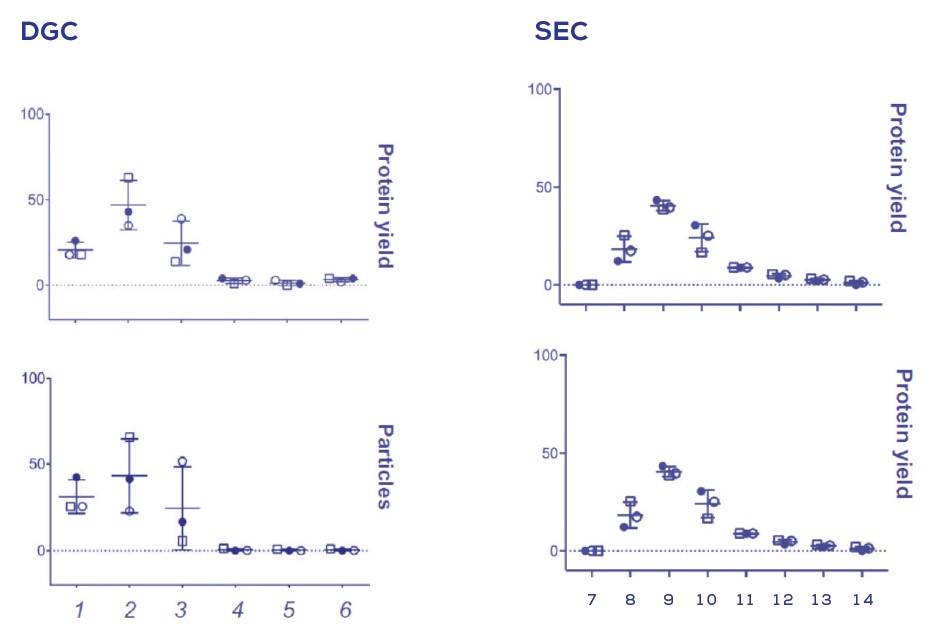
Figure 3. 密度梯度離心(DGC)與尺寸排阻色譜法(SEC)提取bEVs的方法比較
於DGC的密度階層中或SEC-qEV收集體積在已純化的尿路致病性大腸桿菌bEV的粒子與蛋白分佈(佔回收總量的百分比)。
SEC: 一種具有巨大前景與高效的bEV提取工具
Size exclusion chromatography (SEC), 尺寸排阻色譜法, 已被哺乳動物EV研究的團體定位為一種普遍的EV提取方式; SEC的優點在於其簡單而快速的方案、純淨的提取與EV物性/化性的原始程度的保護, 在EV分離時其可標準化的潛力、高重複性和高特異性都是其優勢之一。
儘管在真核生物的EVs分佈中, 以SEC為基礎的bEVs提取還只在最近幾年內開展, 但特別的是, 以SEC為基礎的qEV columns已成為可靠且方便的bEV提取工具; qEV columns的種類可包容不同的處理體積的需求和其奈米粒子提取的範圍。
在Figure 2中, qEV columns顯示出從可溶性蛋白或團聚物中分離bEVs, 分別出現於早先與後面的流洗體積中出現; qEV columns也被拿來與DGC法做對比, 在三重覆的bEV製劑中使用qEV columns可產生更好的重覆性(Figure 3)。
另外, 當使用於bEV純化時, qEV columns和DGC法都在bEV蛋白的消耗與富集展示了可相比的程度; 同時, 在不同的菌種與菌株中, 以超濾法結合qEV提取的濃度可顯示出高效率的大尺度分離和bEV提取的重覆性。
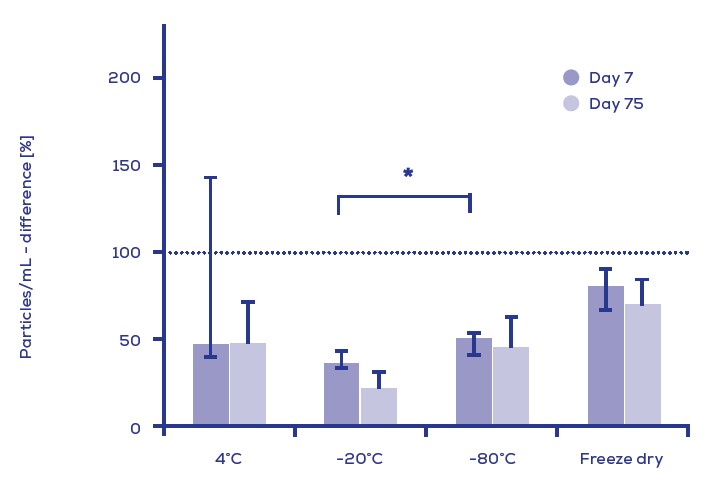
Figure 4. bEV製劑隨著時間與溫度的穩定度
儲存於PBS中黏細菌的bEVs(4度C, -20度C, -80度C或冷凍)於7天與75天後的粒子濃度回收率。
bEV提取後的考量要點
在已經過純化的bEVs中最常使用的緩衝液是PBS, HEPES和低強度的Tris-HCl(20 mW); 關於哺乳動物的EVs的儲存穩定性已被很廣泛的研究了, 且結果可被轉移到bEV製劑中; 例如bEVs也是由磷脂雙層所分隔。
然而細胞壁衍生的成分可能被包含在某些bEV中, 這些成分不會顯示出其對溫度變化的抗性; 以真核生物的EVs為例, 特異性bEVs的研究已顯示出在-80℃保存與冷凍乾燥方式都提供了比在20℃和4℃下保存有更好的回復率(Figure 4)。
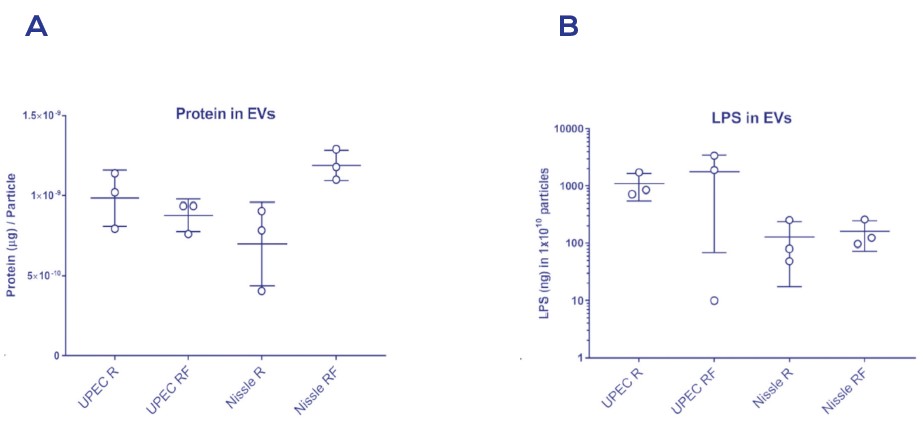
Figure 5. 用低鐵(R)或高鐵(RF)培養的大腸桿菌泌尿致病(UPEC)或益生菌 (Nissle)菌株中以SEC的qEV純化而得的bEVs的分子含量的差異
在bEV製劑中的蛋白含量(A)與LPS含量(B)。
值得注意的是, 從革蘭氏陰性菌中提取的bEVs可自然附著於親代細胞所衍生的內毒素或脂多醣(LPS)分子上; 然而, LPS分子也能以可溶性形式被釋放到細胞外空間, 意識到這種高度具免疫原性分子的存在於bEV的製劑中, 特別對於在功能研究中使用有意義的實驗對照組進行測試是很關鍵的。
Figure 5突出了這個概念, 顯示其在不同的培養條件與相同物種的菌株中, 可於bEV的製劑中發現LPS。
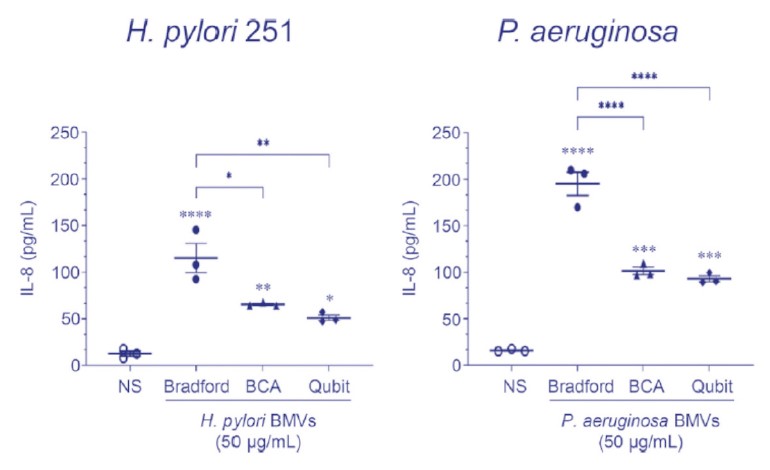
Figure 6. 依照所選擇的bEV蛋白定量測定於下游bEV功能性測定的差異
使用Bradford, Lowry, BCA或Qubit蛋白定量測定法定量從幽門螺桿菌或銅綠假單胞菌用 50 μg/mL bEV處理的AGS細胞中的白細胞介素8的分泌。
在bEV提取中的細胞去除不完全或外部環境污染的潛在風險, 一般都會導致需要對bEV的製劑進行無菌的檢查, 這可採用轉移等分試樣的bEV製劑到培養盤中並觀察細菌的生長狀況; 如果確認為是細菌的污染, 建議是將此bEV的製劑丟棄, 污染的細菌細胞會釋放他們自身的bEVs, 幾乎不可能將其與所要的bEVs做區分, 更多的是物染物分泌的LPS可能會盤旋在bEV製劑內或接續的下游測定中。
EV的分析通常是一個有爭議性的主題, 因其有許多的EV分析工具會標的到不同的EV訊號/參數, 在其靈敏度和特性性的表現中存在有顯著的差異; 總粒子的計數和總蛋白數量通常被考量於應用在表示EV的豐度上, 也可用來使EV的處理標準化。
儘管如此, 近期的bEV研究已經指出在由此產生的生物學數據及其解釋中, 選擇基於蛋白而不是基於粒子的分析測定的影響和差異; 例如當用於處理胃腺癌細胞時(Figure 6.), 由不同的蛋白測定法定量而得的 50 µg/mL的bEV蛋白會導致不同的生物學結果(比如細胞因子的分泌)。
因此在bEV的實驗時, 先了解下游分析中的分析選項、可變性和其影響可以在實驗的計劃中做出更好的決定。
bEV在醫學中的應用
bEVs因為其更容易進行基因操作和其生產具有高度的可擴性, 這就能使其在需要大量製造的應用中佔有許多優勢; 此外, bEVs的製造還得益於與已被建立的微生物培養相關的生物處理技術的出現, 可被察覺的是bEV在人類的醫學中最有希望的應用是其用途為針對關於治療劑、疫苗或藥物傳遞方式等。
研究已表明了細菌益生菌中的bEVs具備有效的免疫調節特質, 在一種已知的益生菌大腸桿菌Nissle菌株的實驗中證明了其bEVs可以保護腸致病性大腸桿菌引起的上皮屏障破壞; 另外Nissle bEVs可在特定菌株的方式下活化樹突細胞(DC), 以誘導DC分化為CD4+ T細胞的特定效應子集, 這可在對抗病原體和解決感染問題方面起到主要作用。
因此, 益生菌衍生的bEVs對於傳遞的療法可擁有顯著的潛力; 例如其可做為飲食的補充劑中的功能性食品成分, 目標在改善腸道微生物組的平衡, 或是幫助緩解腸道的發炎。
另一方面, 基於bEVs的病毒可以是一個強力的候選者來產生針對病原體的先天性和適應性免疫反應, 而沒有相關疾病的風險; 靶向疫苗的病原體可於bEVs原產出(親代細胞), 或經由一個完全不同的過程例如將異源抗原整合到生物工程 bEV中。
在有些案例上, 天然的bEVs可呈遞重要的免疫原性抗原或其具有與親代細胞相似的總體抗原指紋, 因此其在產生基於保護性抗體的反應的用途上與同種細菌相比下是成功的; 在這些天然的bEV病毒中, 腦膜炎奈瑟菌bEV病毒的案例已大大的成功應用在許多國家/地區中。
或者說, 有多種方案可以將異源或外源抗原摻入具有不同抗原呈遞效率的bEV疫苗中, 也可以是誘導的或是引入免疫反應的型態(體液或細胞); 加載抗原的選擇包含bEVs中抗原的表面位置或膜腔內、內源性(細菌中的重組抗原表達)或外源性(在bEV純化後添加)的將抗原加載到bEV中、將抗原加載到bEV中的被動或主動程序等。
當然, 多價疫苗可以使用一種或多種抗原加載方案來產生至bEV的骨架中, 而總體來說, 疫苗已成為預防和控制傳染病的有力的公共衛生手段, 其中bEV疫苗可做為多功能且具有成本效益的平台而成為吸睛的候選對象。
另外bEVs還可以成為絕佳的藥物遞送載體, 一個主要的優點是某些蛋白質重組表達系統已經在細菌中被開發出, 使其可以對已建立的方案進行小幅的修改以摻入感興趣的藥物或分子; bEVs的藥物負載過程可以發生在bEV包裝時或bEV純化後, 就如之前所述的用於基於bEV的疫苗的抗原摻入策略相同。
因其可允許附上標籤以將bEV提取進行功能化或部分靶向以改善體內細胞的傳遞, 修飾bEV表面的方法特別令人感興趣; 研究人員已經舉證了用奈米載體大腸桿菌的bEV來成功讓小鼠腫瘤生長成功的消退, 這些bEV展示了其表面可重組腫瘤靶向抗體HER2(人表皮生長因子受體2)並在其膜腔中展示了通過電轉導入的活性抗癌治療成分: 靶向驅動蛋白紡錘體蛋白(KSP)的小干擾RNA(siRNA)。
或者, bEV還可用於將封裝的抗生素傳遞至感染的細菌細胞, 其是基於bEV對細菌細胞膜的親和力以及bEV保護抗生素免受惡劣環境條件影響的能力; 通過這些方式, 可控的bEV在藥物傳遞方面顯示出了巨大的潛力並可做為針對特定細胞(如癌細胞或細菌細胞)的標靶載體。
總結來說, bEV需要大量而專門的研究來推進這些應用, 同時其也將於克服共同的挑戰後受到獲益, 包含大尺度的bEV製造與提取、移除像是LPS的潛在毒性汙染物、不同批次bEV於分子組成的變化和bEV保值期的穩定性。
相關產品
 外泌體尺寸排阻色譜管柱 qEV columns (SEC) |
 外泌體自動餾份收集機 AFC-V2 |
 單顆粒奈米粒徑/數量濃度/Zeta膜電位分析儀 Exoid Nanoparticle Analyzer (TRPS) |
以上內容非經許可請勿任意複製轉發分享
更多的相關訊息, 歡迎和我們聯絡或來信洽詢: info@normanda.com